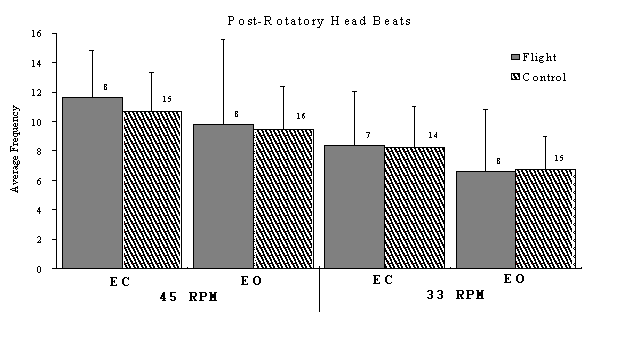
University of Illinois at Chicago,
Department of Electrical Engineering and Computer Science,
Chicago, IL, 60680
R. Kerschmann
University of California at San Francisco
Department of Pathology, San Francisco, CA 94110
R. Sgarioto
Motorola Inc, Desplanes IL 60679
S. Jun
J. Vellinger
SHOT, Floyd Knobs, IN 47119
PUBLISHED: Journal of Vestibular Research, 5: 289-298, 1995.
Robert V. Kenyon, Ph.D
University of Illinois at Chicago
EECS Department (m/c 154)
851 S. Morgan Street
SEO Bldg Rm 1120
Chicago, IL 60607
Office: (312) 996-0450
FAX: (312) 413-0024
kenyon@eecs.uic.edu
Sixty-four fertilized chicken eggs, half at developmental day-2 and half at day-9, were exposed to micro-gravity for 5 days aboard the shuttle. Postflight examination showed that none of the day-2 flight embryos had survived, whereas the day-9 flight group and both groups of synchronous ground control embryos appeared viable. One-half of the day-9 flight and ground control embryos were dissected and the temporal bones preserved in acetone for morphological examination. The other half was allowed to hatch to examine vestibularly related behavioral changes. Morphology of the lagenar otoconia was evaluated by scanning electron microscopy. Behavioral changes were accessed by a battery of reflex tests and recordings of spontaneous and vestibularly driven head movements. The results from both the morphological and behavioral studies showed no consistent difference between the flight and the control animals. Several hypotheses may account for this negative result. Since all the day-2 embryos failed to survive, the remaining day-9 chicks may have passed the critical developmental period of the chick's vestibular system. Also, the re-exposure of the developing chick embryo to earth's 1-g environment may have masked any adverse behavioral effects that exposure to Microgravity may have caused.
Keywords: Spaceflight, Head Movements, Vestibular Morphology, Electron-Microscopy, Chickens, Motor Behavior, Otoconia
In the auditory system, morphological effects have also been produced by sound deprivation. Webster and Webster (13) deprived mice of airborne acoustic stimulation from the age of 3 days and found that these animals had smaller than normal neuron cell bodies in two of the major acoustic brainstem cell groups. Similar results were obtained by Conlee and Parks ( 14) in the auditory magnocellularis nucleus of chicks, and acoustic deprivation has been shown to affect dendritic distribution in another chick brainstem cell group, the nucleus laminaris (15). Such sensory deprivation can lead to permanent behavioral deficits later in adulthood ( 16).
Even normal maturation of the tactile sense has been shown to be dependent on stimulation during development. Simons and Land ( 17) in an electrophysiology study showed that removing the large facial whiskers from rats from the day of birth produced changes in the way individual neurons in the somatosensory cortex responded to stimulation later in life.
While these findings and others may be taken to imply that stimulus dependence is a general feature of a developing sensory system, there are risks in such an extrapolation since each sensory modality has evolved to measure quite different forms of environmental energy. Unlike light and sound, gravity and other external accelerations are transmitted largely unattenuated to the embryo and may therefore play a relatively more fundamental role in embryogenesis. In addition, differences between avian and mammalian species may invalidate such extrapolations across species.
Ground experiments have studied drug effects or the removal of the sensory end-organ early in embryogenesis to try to understand the influence of environmental modifications on the morphology and functioning of the vestibular system. The results of these studies may predict the morphological and behavioral modifications that might be expected should birds develop abnormal vestibular systems from exposure to micro-gravity. In a classic morphology experiment by Levi-Montalcini ( 18) and replicated in a more detailed study by Peusner and Morest ( 19), unilateral extirpation of the prospective inner ear (the otocyst) of the chick at developmental day 2 resulted in abnormal formation of one of the vestibular brainstem nuclei, the nucleus tangentialis.
Decker ( 20) and Heaton ( 21), performing a similar operation, determined that bilateral removal of the primitive ear during the first days of development resulted in serious embryonic behavioral deficits including failure to assume proper position in the egg and consistent failure to hatch, but unilateral extirpation did not significantly alter the behavior of the chick in ovo. However, newborn chicks with only one intact ear showed poor balance and locomotion as indicated by an inability to stand and walk without ataxia. Pronounced abnormalities of head posture were noted, with varying degrees of head tilt, including, in one case, complete 180deg. inversion of the head. The abnormalities were not affected by suppressing visual input. These changes correlated well with morphologic malformations in the operated area and with abnormal microscopic appearance of the ipsilateral nucleus tangentialis, confirming observations of the earlier workers. Heaton noted upon further observation that most of the chicks eventually overcame the reflex deficiencies, and assumed normal behavior and normal head posture within 3-4 days of hatching.
Drugs that damage the vestibular system, for example the antibiotic streptomycin, have also been used to examine vestibular behavior of chicks. Park and Cohen ( 22) injected streptomycin into hatched chicks for several days and showed changes in head stability and body posture. Kenyon et al., ( 23) injected streptomycin into eggs during incubation and showed immediate post hatching effects of vestibular damage both in morphology and behavior. Head instability, first described by Park and Cohen was revealed by Kenyon et al., to be small (5-10deg.) rapid (20 Hz) oscillations that subsided about 7-10 days after hatching. However, this return to normal vestibular behavior was not accompanied by normal morphological structures in the vestibular end organ.
A Soviet experiment flown in earth-orbit aboard the Salyut-4 space station ( 24, 25) showed that frog larval macular hair cells exhibited vacuolization and intercellular edema after 16 days of zero gravity. Otoconia of fish carried on the same mission showed a slight reduction in size, and loss of normal internal concentric and radial lamellation markings. Without inflight centrifuge control data, however, care must be exercised in attributing these changes solely to the removal of gravity. Vacuolization is a non-specific response to stress in many types of cells ( 26). Subsequent orbital studies with frog larvae apparently showed no such changes ( 27, 28 ).
The first attempt at orbiting developing mammals and birds was that of Keefe ( 29), in conjunction with Soviet scientists on board the Cosmos 1129 spacecraft that flew in 1979. Unfortunately, little hard data emerged on morphology and mature vestibular behavior because technical difficulties with the avian portion of the experiment resulted in none of the 60 embryos surviving beyond developmental day 12 of the 20 day mission. The single 10 day old flight embryo that was preserved for study was analyzed by serial light microscopic sections and appeared to be normal.
More recent experiments aboard the Russian MIR space station were successful in hatching 8 quail chicks out of 35 embryos that developed from March 4 through March 24, 1990 ( 30). The hatchlings appeared normal and reacted to sensory stimuli such as vision, touch, and sound. However, the chicks were not able to adapt to the weightless environment. Chicks were unable to feed, or stabilize their bodies by attaching to netting provided for that purpose. The animals were sacrificed after 2-4 days post-hatching and preserved in fixative since death by starvation was inevitable. No results from any follow-up ground studies were available.
The effects of weightlessness on the fully developed inner ear of adult rats was examined in an experiment flown for 7 days on spacelab SL-3. While no evidence of otoconial demineralization was noted in the space-flown animals, an increase in the number of "tiny" otoconia was seen, especially at the border of the utricular endorgan, as compared to control ears. In addition, the otoconia from the saccule showed a more rounded shape in comparison to controls ( 31).
This paper reports on a set of studies of vestibular behavior and otoconial morphology performed on post-flight chicks that were exposed to 5 days of weightlessness aboard Shuttle Discovery in March, 1989 (STS-29).
32 developing chicken embryos (Gallus domesticus) were flown aboard the space shuttle Discovery on a five day mission in March, 1989 (STS-29) ( 32). At launch, the flight and a matched ground control group contained 32 eggs each: half at day 2 and half at day 9 of the 21 day period of development. On return to Earth, one-half of each age group was opened within 2 hours of landing. At this point it was discovered that none of the day-2 flight embryos had survived, whereas the day-9 flight group and all ground control embryos appeared viable. The material from the day-2 flight eggs had undergone extensive autolysis and was deemed unworthy of microscopic examination. Therefore, one-half respectively of the day-9 flight and ground control embryos were dissected and the temporal bones preserved in acetone for later processing for scanning electron microscopy, as described below. Acetone was used for fixation instead of aldehyde-based agents because of the lability of otoconia in certain aqueous solutions. Tissues were also preserved for morphologic study of the sensory epithelium, and will be reported by other authors. The remaining eggs were incubated and after the 21 day developmental period all surviving chicks hatched. Both flight and control chicks were then subjected to a battery of behavioral and vestibulosensory tests to determine the effects of five days of development in Microgravity. The analysis was double-blinded, with random codes assigned to the embryos.
Behavioral tests were administered to chicks between 30 and 60 minutes after hatching. The battery of tests took 20 minutes to administer and included head righting reflexes, post-rotatory head nystagmus and spontaneous head movements. Chicks removed from the incubator for testing were immediately placed in a surgical glove to prevent heat loss during the testing and to restrain their movements. The middle finger of the glove was removed so that the chick's head protruded from this opening, permitting head movements in all directions. The body of the chick was then wrapped in a cloth to further retain heat. Tests were performed with and without vision. A small aluminum foil hood over the chick's head was used to block vision.
The head righting reflexes were conducted by holding the chick by the trunk and tilting it 90deg. to the left, right, forward, back and then inverting the bird. The experimenter noted the ability of the chick to align its head vertically during the rotation and at the subsequent angle of tilt over a period of 10 seconds. Birds with vestibular damage have demonstrated significant changes in head righting reflexes compared with controls (23).
Post-rotatory vestibular nystagmus was stimulated by rotating the bird at 33 and 45 rpm in the clockwise direction for 45 seconds and then suddenly stopping the rotation. The animal was secured in a Styrofoam cup with its head free to move and placed in the center of a turntable. The strength of the resulting head nystagmus was then recorded by counting the number of beats of the head during this post-rotatory period.
Spontaneous head movements were measured to ensure that some subtle vestibularly related head posture abnormality, such as the characteristic head tremor in streptomycin intoxicated chicks (23), would not escape detection. Dynamic spontaneous head movements were recorded using a magnetic search coil system1 (23). The coil system had a sensitively of 260mv per degree and a noise level of 20mv (0.08deg.) and a bandwidth of 250Hz. The output from the coil system was digitized at a sample rate of 1000 Hz per channel (horizontal and vertical) and stored in a computer file for later analysis. The chicks were instrumented by attaching a 20 turn coil of #30 magnet wire to the beak of the bird using paraffin wax (23). The bird was placed in a Styrofoam cup located in the center of the field coils. The chick's attention was then attracted to various positions in the room to stimulate large and small horizontal and vertical head movements. Ninety to 120 seconds of recordings were made per animal.
Analysis
Post-rotatory head beats were statistically analyzed by averaging data from all control populations according to test condition. In addition, day-2 and day-9 control data were separated and compared to each other and to day-9 flight chick data.
Dynamic head movements were analyzed by calculating the velocity of the movements from the position data using a 3-point difference algorithm. This gave the original 1000 Hz sampled data a velocity bandwidth of 200 Hz. Head position and velocity were displayed simultaneously using a computer and the velocity and amplitude extracted from the data using a digital cursor. The resulting peak velocity and amplitude data were then plotted on a log-log plot.
Scanning Electron Microscopy
Tissue from one ear of three flight and five ground control animals was available for analysis by scanning electron microscopy (SEM). After transportation, temporal bones were further dissected in a pool of acetone under a dissection microscope. Lagenar otolithic membranes were teased free of the bone and mounted on standard SEM pedistals and the acetone allowed to evaporate. The specimens were then gold-palladium sputter coated in a model Hummer X specimen coater (Anatech Corp.) and viewed and photographed in a JSM-480 (JEOL) scanning electron microscope. The small number of available flight-animal ears precluded any dependable statistical analysis of the data.
The behavioral tests showed no consistent difference between the flight-chicks and the ground-control populations. Righting reflexes appeared normal in the flight population with and without the hood. Chicks could maintain their head aligned with the vertical at any orientation and changing the speed at which the body was tilted did not reveal any abnormal or sluggish reflex responses.
Figure 1. The average number of head beats of post-rotatory nystagmus for flight and control populations for two rotation speeds (33 and 45 rpm) with eyes covered (EC) and eye open (EO) conditions. Sample size is indicated by the number next to the standard deviation error bar for each condition.
The post-rotatory nystagmus response never exceeded 10 seconds in duration for either flight or control animals. The data in Figure 1 illustrates the close similarity of the responses recorded from each group. None of the differences between the populations shown in this figure are significant. In general, the data shows that the higher rotation velocity yielded higher number of post rotatory beats. In addition, the eyes covered (EC) condition had a higher number of beats than eyes open (EO).
(A)
(B)
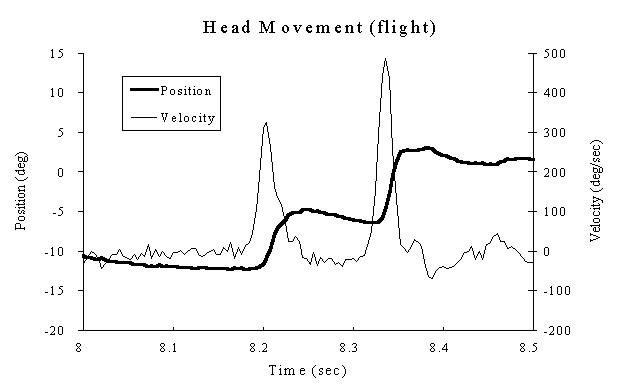
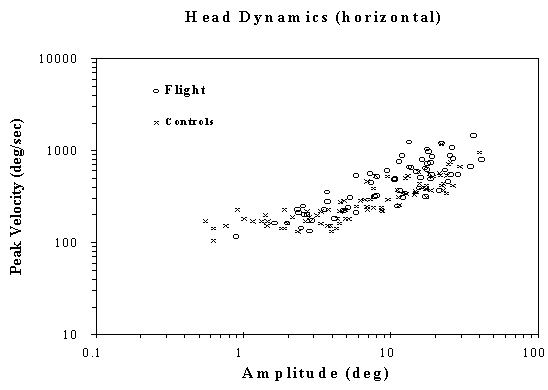
Figure 2. (a) A typical (magnetic coil system) recording of spontaneous head movements (heavy line; left Y-axis) and its related velocity (light line; right Y-axis) from a flight chick. Control of head position between movements were stable and unremarkable. (b) Relationship between head movement amplitude and peak velocity for ground-control (plotted as "x") and flight populations (plotted as "o"). Both populations show the same relationship between position and peak velocity.
Recordings of spontaneous head movements using the magnetic search coil showed that head position was stable between movements without any tremor or oscillations, as shown in the flight animal recording in Figure 2a. Chick head movement dynamics, Figure 2b, plotted as head peak velocity verses head amplitude, shows that data from each population are intermingled and do not show any significant differences in this dynamic characteristic. Each set of data shows a rise in peak velocity as head amplitude increased above 4deg. with the data below this leveling off at 100deg./sec.
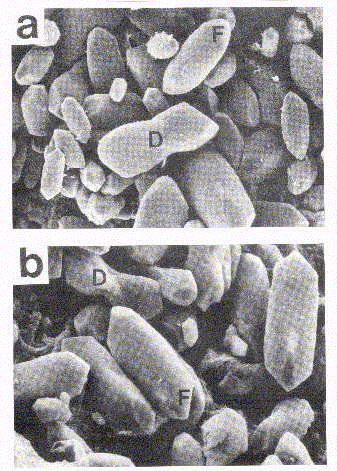
Figure 3. Scanning electron microscopy of lagenar otoconia from 14 day flight (a) and vibration control (b) embryos. The otoconial morphology is typical, with barrel-shaped profile and triple faceted ends (F) evident in both groups. Normally occurring variants, including double fluted forms (D) were also present. (scale, 1cm = 2.2).
Lagenar otolithic membranes showed no significant abnormalities as examined under SEM. Both flight (Figure 3a) and control (Figure 3b) material showed the usual barrel-shaped otoconial profile with triple-faceted ends. Scattered among these unremarkable otoconia were occasional immature forms, including so called double-fluted types ( 33), as well as crossed otoconia.
The absence of significant and consistent changes in behavior and morphology between the flight and control chicks in our study does not necessarily indicate that micro-gravity is a benign environment for chick vestibular development. For example, all day-2 embryos on this experiment were lost and only 8 of 35 quail embryos succeeded in hatching in the Russian experiment (30). In addition, a recent experiment on board Spacelab-J (STS-47) found that only 1 of 15 day-0 embryos survived mission ( 34). The loss of the day-0 embryos is believed to be due to failure of the yolk sack to emerge from the albumin due to specific gravity differences between the two structures ( 35).
In terms of possible vestibular modifications due to abnormal development, the loss of the day-2 embryos is significant because the vestibular system has not formed in these animals whereas in a day-9 embryo the vestibular system is developed to such an extent that vestibular reflexes are observable ( 36). In general, otolith formation starts on day 4 ( 37), and sensory connections between vestibular sensory epithelium and the brainstem are initiated on day 5 ( 38). Consequently, it is likely that the vestibular system has passed its "critical period" by day 9. While a postnatal "critical period" has been shown to exist in other sensory systems for several weeks to months, the nature of any similar period of susceptibility in the vestibular system is unknown.
An exposure to microgravity of 5 days may not be long enough to produce significant changes in vestibular function. In many of the sensory modification experiments done on other systems, a length of exposure of at least several weeks was needed to show effects of the modified environment when the critical period was unknown (8). In addition, in such studies the stimulation of the sensory system was meticulously controlled at all times so that unwanted stimuli were removed. In the present experiment, the length of exposure was not controllable nor was the endogenous and exogenous acceleration stimuli experienced by the embryo completely eliminated. As the embryo develops, somatic muscle contractions take place in the embryo. Normally, spontaneous head flexions produced by chick embryos first appear about days 3-4 of incubation, followed by distinct movements of the trunk at 5 days ( 39). These motions are cyclic, with 5-15 second activity periods separated by 30-60 second intervals of immobility. Such spontaneous movements on the part of the embryo produce both angular and linear accelerational forces at the location of the developing inner ear, constituting a major source of transient stimulation of the vestibular system. In addition, rhythmic 15-20 per minute contractions arise de novo in the smooth muscle of the amnionic membrane ( 40), which functions independently of the embryo proper ( 41). These endogenous accelerations along with those imposed on the embryo by the accelerations of the orbiter constitute a level of stimulation that may be sufficient for normal vestibular development.
Despite the presence of transient accelerations, chronic exposure to micro-gravity might have produced changes in the vestibular system initially which could have been masked by the adaptation of the chick to the reassertion of the 1-g field for 7 days postflight prior to hatching. Previous experiments on chicks that have been severely impaired by various modifications applied to the vestibular system have shown a remarkable ability to behave normally after a few days to a week of adaptation. Decker (20) and Heaton (21) showed that newly hatched chicks with only one intact ear and vestibular response deficits eventually overcame the reflex deficiencies, and assumed normal behavior and normal head posture within 3-4 days of hatching. Park and Cohen (22) and Kenyon et al. (23) found that small rapid head oscillations from streptomycin-induced damage of the vestibular system in chicks subsided after about 7-10 days as did the abnormal head reflexes and postural ataxia. However, this return to normal vestibular behavior was not accompanied by re-establishment of normal morphological structures in the vestibular end organ of the streptomycin chicks.
Given the rapid adaptation of the chicks after such drastic deficits of vestibular function, it might be necessary for chicks to be studied in space. For example, the Russian experiments showed that quail embryos can hatch successfully in weightlessness, but the chicks were not able to adapt to weightlessness over a period of 2-4 days. However, the normalcy of their vestibular structures and behavior is still a question. Perhaps, more sensitive behavioral tests might have uncovered changes that existed only immediately post-hatching. For example, Jones et al.,( 42), showed increased vestibular thresholds in this same population of flight animals some 21 days post-hatching. Unfortunately, the lack more sophisticated equipment and the short time available for post-flight testing limited these experiments.
The SEM data presented here show that there was no gross effect of micro-gravity on the vestibular otoconia morphology of newly hatched chickens. While the utricular and saccular otoconia were not available for scanning electron microscopy, we examined a small number of specimens for irregularities of the lagenar otoconia and found them to be normal in the flight embryos. Some differences in the size of saccular otoconia in space flown rats has been described ( 43), systematic difference in otoconial size between the experimental and control groups in our material was not identified; what small variation was seen could be attributed to sampling error across the membrane, since the stones nearer the edges to the organs are known to be smaller than those more centrally located (43). Studies to examine the morphologic details of the vestibular sensory cells themselves are underway, and will be reported by a separate group of scientists.
The authors thank Kentucky Fried Chicken Corp. (KFC) for supporting this research, the staff at KFC Headquarters in Louisville, KY, for their hard work and assistance with our experiments, and the members of the science team, Ron Hullenger and Scotty Hester, for their advice and help with the post-flight work. This work was supported in part by UIC Campus Research Board grant #87-402 awarded to R. Kenyon.
1. Sosula L, Glow PH. Increase in number of synapses in the inner plexiform layer of light-deprived rat retinae: quantitative electron microscopy. J. Comp. Neurol. 1971; 141: 427-452.
2. Gyllensten L, Malmfors T. Myelinization of the optic nerve and its dependence on visual function - a quantitative investigation in mice. J. Embryol. Exp. Morph. 1963; 11, part 1: 255-266.
3. Wiesel T, Hubel D. Effects of visual deprivation on morphology and physiology of cells in the cat's lateral geniculate body, J. Neurophysiol. 1963a; 26: 979-993.
4. Wiesel T, Hubel D. Single cell response in striate cortex of kittens deprived of vision in one eye, J. Neurophysiol. 1963b; 26: 1003-1017.
5. Wiesel T, Hubel D. Comparison of the effects of unilateral and bilateral eye closure on cortical unit responses in kittens, J. Neurophysiol. 1965a; 28: 1029-1040.
6. Lund JS, Lund RD. Development of synaptic patterns in the superior colliculus of the rat. Brain Research 1972; 42: 1-20.
7. Raviola E, Wiesel TN. An animal model of myopia. New England Journal of Medicine 1985; 312:1609-1615.
8. Hubel D, Wiesel T. The period of susceptibility to the physiological effects of unilateral eye closure in kittens. J. Physiol. 1970; 206: 419-436.
9. Wiesel T, Hubel D. Extent of recovery from effects of visual deprivation in kittens, J. Neurophysiol. 1965b; 28: 1060-1072.
10. Dews P, Wiesel T. Consequences of monocular deprivation on visual behavior in kittens, J. Physiol. 1970; 206: 437-455.
11. von Noorden G. Histological studies of the visual system in monkeys with experimental amblyopia, Invest. Ophthamol. 1973; 12: 727-738.
12. Ikeda H, Wright M. Properties of LGN Cells in kittens reared with convergent squint: a neurophysiological demonstration of amblyopia, Exp. Brain Res. 1976; 25: 63-77.
13. Webster, DB, Webster, M. Neonatal sound deprivation affects brainstem auditory nuclei. Arch. Otolaryngol 1977; 103, 392-396.
14. Conlee JW, Parks TN: Age-and position-dependent effects of monaural acoustic deprivation in nucleus magnocellularis of the chicken. J. Comp. Neurol. 1981; 202, 373-384.
15. Gray L, Smith Z, Rubel EW: Developmental and experimental changes in dendritic symmetry in nucleus laminaris of the chick, Brain Res. 1982; 244: 360-364.
16. Tees RC: Effects of early auditory restriction in the rat on adult pattern discrimination. J. Comp. Physiol. Psychol. 1967, 63, 384-393.
17. Simons DJ, Land PW: Early experience of tactile stimulation influences organization of somatic sensory cortex. Nature 1987; 326:694-697.
18. Levi-Montalcini R. The development of the acoustico-vestibular centers in the chick embryo in the absence of the afferent root fibers and of descending fiber tracts. J. Comp. Neurol. 1949; 91: 209-241.
19. Peusner KD, Morest DK. Neurogenesis in the nucleus vestibularis tangentialis of the chick embryo in the absence of the primary afferent fibers. Neuroscience 1977; 2: 253-270.
20. Decker JD. The influences of early extirpation of the otocysts on development of behavior in the chick. J. Exp. Zool. 1970; 174: 349-369.
21. Heaton MB. Behavioral adaptation in neonatal chicks following embryonic vestibular system rearrangement. J. Exp. Zool. 1975; 194: 495-510.
22. Park JC, Cohen GM. Vestibular ototoxicity in the chick: effects of Streptomycin on equilibrium and on ampullary dark cells. Am J Otolaryngol 1982; 3: 117-127.
23. Kenyon R, Kerschmann R, Silbergleit R. Streptomycin in the chick embryo: post-hatching vestibular behavior and morphology. Exp. Brain Res. 1988; 69: 260-271.
24 Vinnikov Y. The role of gravity in the phylogeny of structure and function in animal sensors of spatial orientation, and their predicted action in weightlessness. Life Sciences and Space Research XII 1974; 59-176.
25 Vinnikov Y. The development of the vestibular apparatus under conditions of weightlessness. Arkhiv/anatomii, gistologii i embyriologii LXX 1976; No. 1, Washington NASA TT F-16987.
26 Robbins S, Cotran R. Pathologic Basis of Disease 2nd ed. W.B. Sanders Co. 1979.
27 Young R, Tremor J. Weightlessness and the developing frog egg. Life Sciences and Space Research 1968; 6: 87-93.
28 Vinnikov Y, Gazunko OG, Lychakov DV, Palmbakh CR. The development of vestibular apparatus under the conditions of weightlessness. Zhurnal, Obshchei. Biologii 1983; 44: 147-163.
29. Keefe JR. Experiment K-313: Rat and quail autogenesis. In Final Reports of U.S. Rat Experiments Flown on the Soviet Satellite Cosmos 1129 NASA TM-81289, 1979.
30. Meleshko G, Shepelev E, Gurieva T, Bodya K, Sabo V. Embryonic development of birds in weightlessness. Space Biol. and Aerospace Med. 1991; 25, 37: 37-9 (transl. from Russian by P.V. Chetirkin).
31. Ross MD, Donovan K, Chee O. Otoconial morphology in space-flown rats. The Physiologist 1985; 28: S-219-220.
32. Vellinger J, Deuser M. Avian embryogenesis in microgravity aboard Shuttle STS-29: Experimental protocol and results. Am. Soc. Grav. Space Biol. Bulletin 1990; 4(1): 74.
33. Ballarino J, Hawland H. Otoconial morphology of the developing chick. Anatom. Record 1982; 204: 83-7.
43. Ross MD, Donovan K. Otoconia as test masses in biological accelerometers: what can we learn about their formation from evolutionary studies and from work in micro-gravity? Scanning Electron Microscopy 1986; 4: 1695-1704.
34. Suda T. STS-47/Spacelab-J: Life science investigations. Am. Soc. Grav. Space Biol. Bulletin 1993; 9(1): 13.
35. Suda T, Abe E, Shinki T, Katagrir T, Yamaguchi A, Yokose S, Yoshiki S, Horikawa H, Cohen G, Yasugi S, Naito M. The role of gravity in chick embryogenesis. FEBS Letters 1994; 340: 34-38.
36. Gottlieb G. Prenatal behavior of birds. Quart. Rev. Biol. 1968; 43: 148-174.
37. Fermin CD, Igarashi M. Otoconia formation in the chick embryo. In Proceedings of the 41st Annual Meeting of the Electron Microscopy Society of America, G.W. Baily, ed., 1983.
38. Knowlton VY. Correlation of the development of membranous and bony labyrinths, acoustic ganglia, nerves, and brain centers of the chick embryo. J. Morph. 1967; 121: 179-202.
39. Hamburger V, Balaban M. Observations and experiments on spontaneous rhythmical behavior in the chick embryo, Dev. Biol. 1963; 7: 535-545.
40. Pierce ME. The amnion of the chick as an independent effector. J. Exp. Zool. 1933; 65: 443-473.
41. Oppenheim RW. Amnionic contraction and embryonic motility in the chick embryo. Science 1966; 152: 528-9.
42. Jones T, Vellinger J, Hester P, Fermin C. Weightlessness and the otogeny of vestibular function: Evidence for persistent vestibular threshold shifts in chicks incubated in space. The Physiologist (abstract) 1992; 34 (supl. 1): 5143-5.